

Одним из самых важных органов чувств считаются глаза. Благодаря им мы получаем львиную долю информации об окружающем мире. Когда мы рассматриваем что-то, наш взор не фиксированный, т.е. наши глаза совершают множество мелких движений, именуемых саккадами. Из-за этих движений смещается сетчатка, что в свою очередь снижает четкость изображения (вспомните не самые удачные фильмы в стиле «мокьюментари»). Чтобы компенсировать этот эффект, зрительная система снижает свою чувствительность к смещению зрительных стимулов во время саккад. Однако до сего времени механизмы, лежащие в основе этого тонкого процесса, оставались не изучены. Посему ученые из университета Тохоку (Япония) решили провести ряд практических опытов и моделирований. Что показали эксперименты, какие секреты таит в себе гашение саккадического смещения, и почему так важны визуальные стимулы до и после саккад? Ответы на эти вопросы мы найдем в докладе ученых. Поехали.
Основа исследования
Глаза человека, как мы знаем, не самые лучшие в сравнении с другими животными на нашей планете. Однако их нам вполне хватает, чтобы получать информацию об окружающем мире.
Важнейшей частью зрительной системы, помимо центральной нервной системы, является сетчатка глаза. В ней содержатся фоторецепторы, которые воспринимают электромагнитное излучение видимого спектра и преобразуют его в нервные импульсы. Последние поступают в мозг, где и формируется картинка, которую увидели глаза.
Глаза человека являются одним из важнейших органов для диагностики множества заболеваний, которые могут быть даже не связаны со зрительной системой.
Чего не хватает сетчатке, так это однородности: большая часть зрительного поля связана с периферическим зрением и лишь небольшая центральная часть отвечает за основное (сфокусированное) зрение. Разница между этими областями очевидна: периферическое зрение имеет меньшее пространственное разрешение, чем центральное. Следовательно, чтобы эффективно собирать визуальную информацию, глаза должны двигаться.
Эти движения, как уже говорилось ранее, называют саккадами и совершаются глазом несколько раз в секунду. За счет саккад мы меняем точку фиксации нашего взгляда, за счет его осуществляется рассматривание объекта интереса, т.е. сбор визуальной информации.
Саккады работают во всех направлениях (вверх-вниз, влево-вправо), однако наибольшее число движений происходит в горизонтальной плоскости. А диапазон саккад варьируется от 2’ до 15°.
Саккады во время поворота глаза.
Совокупность малого центрального поля зрения и саккад приводит к появлению определенных проблем. Во-первых, изображение сетчатки становится размытым во время саккады, и это может ухудшить зрительное восприятие. Во-вторых, по всей саккаде положение объекта на сетчатке меняется. Это приводит к значительному изменению изображения на сетчатке, что в свою очередь приводит к осложнениям во время «расшифровки» мозгом изображений до и после саккады.
Ученые напоминают нам, что визуальная чувствительность глаз уменьшается во время саккад, если происходят определенные изменения с объектом рассмотрения (изменение освещения, смещение объекта, постоянное движение объекта, изменение цвета и тд). Это явление называется саккадическим подавлением или саккадическим упущением путем прямой и/или обратной маскировки.
Саккадическое подавление может препятствовать нормальному восприятию размытого изображения сетчатки, размытость которого вызвана именно саккадой.
Вторую проблему, связанную со смещением проекций объектов на сетчатку, труднее решить, поэтому возникает проблема интеграции изображений сетчатки до и после саккады. Существует теория, что удаления изображения во время саккад, т.е. останутся только изображения до и после, может способствовать стабилизации визуальной информации. Однако это не так, поскольку отличия в изображениях до и после все равно остаются.
Потому ученые и предполагают, что механизм оценки саккад-индуцированного сдвига в изображении на сетчатке основан на копии эфферентной* моторной команды движения глаза для компенсации сдвигов изображения.
Эфферентные* (двигательные) нейроны передают импульс ЦНС рабочим органам.После перенастройки местоположения стимула в пространстве механизм может интегрировать изображения сетчатки до и после саккад. Тем не менее, поскольку фактическая амплитуда саккады и амплитуда саккады, предсказанная на основе эфферентной копии, не всегда одинаковы, объект может восприниматься как смещенный во время саккады даже без физического смещения, если визуальная система полностью доверяет копии.
Решение этой проблемы заключается в том, что зрительная система намеренно не фиксирует небольшие смещения объекта во время саккады, даже если их легко обнаружить во время фиксированного рассмотрения. Этот феномен называют саккадным подавлением смещения (SSD от saccadic suppression of displacement).
Наличие определенного количества допуска (допустимой неточности, погрешности или ошибки) к смещению объекта через саккады может быть простым способом достижения эффективной стабильности визуального восприятия.
Еще одним важным элементом в исследовании визуальной стабильности является эффект гашения (blanking effect).
Эффект гашения — это явление, при котором эффективность обнаружения смещения объекта во время саккады улучшается за счет вставки временного пробела (50–300 мс) во время саккады и до того, как объект вновь появляется в новой позиции. Это явление предполагает, что сигналы о смещении объекта действительно доступны после саккады, но не могут использоваться для обнаружения смещения, когда объект саккады присутствует в ее конце, т.е. без пост-саккадического пробела. Смещение, однако, может быть воспринято, когда подавление устраняется путем вставки пост-саккадического пробела.
Эффект гашения поддерживает предположение о том, что SSD является активным процессом, позволяющим избежать любого ошибочного восприятия смещения, вызванного неточными саккадами и/или неточными копиями эфферентных команд саккады, поскольку оно демонстрирует, что имеется достаточно информации для обнаружения смещения после саккад.
В ранее проведенных исследованиях было установлено, что магноцеллюлярный зрительный путь (М-путь), который реагирует на стимулирующее движение, избирательно подавляется во время саккад, как и низкоуровневые движения. Гашение или удаление объекта сразу после саккады активирует механизм, который подавляет М-путь во время саккады.
Также считается, что важную роль в механизме саккад играет контрастность объекта рассмотрения. Чтобы проверить это, ученые провели анализ влияния контраста до- и после-саккадического обэекта на обнаружение смещения.
Поскольку время демонстрации объекта и его положение до саккады было одинаковым как в обычных условиях, так и при использовании эффекта гашения, любое различие в обнаружении должно быть связано с объектом после саккады.
Было установлено, что обнаружение смещения было лучше, если объект до саккады имел более высокую контрастность. А вот увеличение контрастности объекта после саккады приводило к ухудшению восприятия в обычных условиях, но не в случае применения эффекта гашения.
Подобные наблюдения говорят о том, что более сильный сигнал от объекта после саккады активирует механизм обнаружения смещения объекта.
Подготовка к опытам
Однако этот анализ является лишь частичным объяснением того, как работают саккады и сопряженные с ними механизмы восприятия визуальной информации. Для более точного изучения были проведены практические опыты с добровольцами. После чего также была создана модель, которая объясняет что и как работает.
Участниками наблюдений стали 8 мужчин и 2 женщины в возрасте от 21 до 24 лет с нормальным зрением. Каждый из участников размещал голову на специальной подставке (как в кабинете офтальмолога) на расстоянии 45 см от объекта, демонстрируемого на ЭЛП-экране (Sony GDM-F520, 21″, 100 Гц).
Положение глаз измерялось с помощью устройства слежения за лимбом*, состоящего из инфракрасного диода и двух фотодиодов.
Лимб роговицы* — место соединения роговицы и склеры, т.е. край роговицы.Положение глаз и скорость их движения оценивались в реальном времени посредством алгоритма разности трех точек, в котором скорость в момент времени n рассчитывалась исходя из положения глаза в момент времени n-1 и момент времени n+1.
Начало саккады определялось как момент, когда скорость глаза превышала 30 °/с. Обнаружение начала саккады всегда задерживалось на 1 кадр (10 мс) из-за метода расчета скорости.

Условия опытов: А — опыты без применения временных пробелов; В — опыты с временными пробелами (эффект гашения).
Одна из основных сложностей подобных экспериментов, как заявляют сами ученые, заключается в правильном тайминге всех этапов. Существует вероятность, что участник опыта может увидеть исчезновения предсаккадного объекта только по завершению саккады. А это может привести к искусственному улучшению восприятия смещения. Потому был проведен точный расчет времени смещения саккады и времени демонстрации объекта.
Расчеты показали, что исчезновение предсаккадического объекта происходит примерно за 32 мс до самой саккады. Лишь в 0.7% опытов объект исчезал после саккады. Также в дальнейшем моделировании не учитывались результаты опытов, где саккада была короче 120 мс или дольше 400 мс. Суммируя все уточнения и исключения, было забраковано лишь 6% результатов опытов.
Также необходимо было провести калибровку перед каждым опытом. В начале калибровки пять одинаково разделенных точек были представлены последовательно вдоль горизонтальной линии, тогда как центральная точка была всегда расположена в центре дисплея. Участникам необходимо было фиксироваться на каждой точке по очереди и нажимать кнопку, когда каждая фиксация была завершена. Если коэффициент регрессии* был меньше 0.9, процедуру калибровки повторяли до тех пор, пока критерий не был удовлетворен.
Коэффициент регрессии* показывает силу и характер влияния независимых переменных (в данном случае 5 точек) на зависимую.В качестве объектов (точка фиксации взгляда, объект до и после саккады), за которыми должны были наблюдать участники опытов, выступали диски диаметром 0.88°. Фоновая яркость была постоянной и составляла 21.4 кд/м2 (кандел на метр квадратный). Перед проведением опытов также был измерен контраст. В данном случае это был контраст Вебера, который является отношением объекта к фону, и поэтому в некоторых случаях контраст превышал 100%. Чтобы учесть разницу в чувствительности между центральным и периферическим зрением, контрасты стимула определялись факторами порогового контраста.
Контраст до-саккадического объекта в 3, 4.3, 6.1, 8.8, 12.6 или 18 раз превышал порог контраста при периферическом зрении, а у пост-саккадического — в 3, 4.3, 6.1, 8.8, 12.6 или 18 раз превышал порог контрастности при центральном зрении.
Результаты опытов и моделирования
Первым делом стоит определить, каково же влияние контраста на обнаружение смещения.
Сначала участникам опытов необходимо было выполнить саккаду вправо под углом 17.8° от точки фиксации взгляда до объекта (диска). Как только начиналась саккада, объект в первоначальной позиции исчезал и появлялся на 0.33° вправо или влево сразу (без временно пробела) или с задержкой в 100 мс (эффект гашения). Основная задача участников — сказать куда сместился объект (влево или вправо). Значение контраста объекта до и после сакадды варьировалось в ходе опытов.

Изображение №1: влияние контраста до и после саккады на точность обнаружения смещения при обычных условиях (без эффект гашения).
Точность обнаружения смещения объекта через саккаду выражалась как d в каждом контрастном состоянии. Значения d в условиях отсутствия/наличия временных пробелов показаны на изображениях 1 и 2 соответственно.

Изображение №2: влияние контраста до и после саккады на точность обнаружения смещения при использовании временного пробела (эффект гашения).
В состоянии без временного пробела точность обнаружения смещения значительно снизилась с постсаккадическим контрастом (1C), а с досаккадическим контрастом наоборот улучшилась (1А).
В режиме с временным пробелом точность обнаружения смещения улучшалась при контрасте до саккады (2А), но никак не менялась от контраста после саккады (2С).
Факт того, что повышение контраста до саккады улучшает обнаружение смещения объекта, полностью согласуется с классическим представлением о работе глаз человека: чем сильнее первичный стимул, тем лучше будет его дальнейшее обнаружение. А вот снижение точности глаз из-за повышения контраста объекта после саккады подразумевает, что постсаккадический стимул усиливает SSD (саккадное подавление смещения).
Для лучшего понимания связи между SSD и контрастности ученые провели моделирование.
Информация от сетчатки передается в зрительную кору по двум основным функционально отличным путям: магноцеллюлярный путь (M-путь) и парвоцеллюлярный путь (P-путь). Психофизические исследования показали, что M-путь сильно подавлен во время саккад.
Получается, что информация о яркости объекта, представленного сразу после саккады, сильно влияет на эффективность обнаружения смещения, как если бы подавление M-пути было еще больше усилено увеличением контрастности.
Если М-путь подавлен во время саккад, то Р-путь должен быть доминантным сразу после саккад. Это значит, что постсаккадическая информация от сетчатки обрабатывается главным образом через P-путь. Снижение точности с увеличением постсаккадического контраста наталкивает на мысль, что активность P-пути подавляет обнаружение смещения.
Чтобы проверить эту теорию, ученые создали модель, основанную на предположении, что P-путь доминирует сразу после саккады.
Если предположить, что существует только два визуальных пути от сетчатки до коры (игнорируя все остальные), одна из самых простых моделей будет состоять в том, что сигналы M-пути подавляются сигналами из P-пути или визуальных областей более высокого уровня, которые получают входной сигнал от P-пути.
Отклик каждого пути на контраст был выражен функцией, которая также используется для моделирования контрастного ответа ЛКТ* клеток М- и Р-типа и для сравнения с психофизическими результатами опытов.
ЛКТ* (латеральное коленчатое тело) — структурная часть мозга, расположенная на нижней латеральной стороне подушки таламуса.Для оценки выходного сигнала обнаружения смещения была использована модель Рейхардта, с помощью которой была рассчитана корреляция между пред- и постсаккадическими сигналами, косвенно связанными с M-путем.
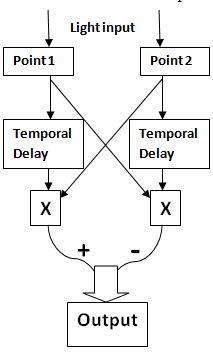
Модель Рейхардта для оценки обнаружения смещения объекта.
Если утрировано, то в данной модели от значения отклика Р-пути зависела сила выходного сигнала всей модели (чем выше отклик Р-пути, тем ниже отклик всей модели).
Следующая формула вычисляет выходные данные модели, используемой для выражения влияния контраста до и после саккады на точность обнаружения смещения:
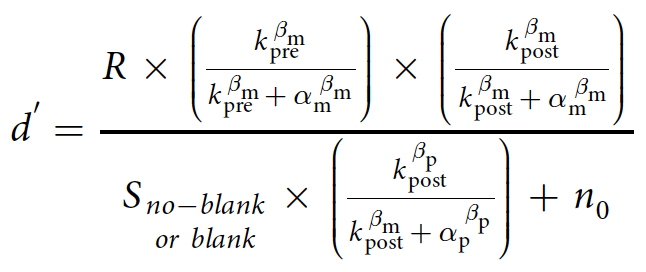
k pre — нормализованный контраст объекта до саккады;
kpost — нормализованный контраст объекта после саккады;
параметры α и β — контраст полунасыщения и сила зависимости от контраста;
R — коэффициент, используемый для преобразования отклика в точность обнаружения смещения;
n0 — константная нелинейная функция, моделирующая нейронные контрастные характеристики, которая контролирует уровень шума;
m и p — M-путь и P-путь;
S — вклад отклика P-пути, который влияет на обнаружение смещения;
SBlank — вклад отклика P-пути в случае использование эффекта гашения (т.е. временного пробела).

Таблица №1: подогнанные параметры модели. Числа показывают параметры, полученные путем подгонки предлагаемой модели к данным точности обнаружения смещения, усредненным по всем участникам опытов.

Изображение №3: результаты моделирования.
Согласно таблице №1, функция P-пути имеет более низкий контрастный отклик и меньший контраст полунасыщения, чем функция M-пути, что соответствует разнице в α (αp в 2.4 раза больше, чем αm). Это согласуется с разницей между характеристиками контрастного отклика ЛКТ клеток P- и M-типа, о которых сообщалось в ранее проведенных исследованиях: контраст полунасыщения ЛКТ клеток P-типа приблизительно в 5 раз больше, чем у клеток M-типа.
Результаты моделирования полностью согласуются с гипотезой о разном влиянии двух путей на обнаружение смещения. Таким образом, Р-путь вносит положительный вклад в саккадное подавление смещения, а М-путь способствует обнаружению смещения объекта.
Для более подробного ознакомления с нюансами исследования рекомендую заглянуть в доклад ученых и дополнительные материалы к нему.
Эпилог
В данном труде ученые установили, что именно информация от сетчатки, полученная сразу после саккады, активирует механизм обнаружений смещения. Это означает, что пост-саккадическая информация сетчатки является важным фактором в отношении зрительной стабильности.
Ученые отмечают, что существует две основные теории, описывающие стабилизацию зрения. В первой теории зрительная система преобразует информацию о положении пре-саккадической цели в пост-саккадические координаты. За счет переназначения позиции «до» и «после» может быть выполнено сравнение позиций в пространстве, таким образом компенсируя смещения сетчатки, вызванные саккадой. Одной из основных тонкостей этой теории является гипотеза, что смещения сетчатки, вызванные саккадами, могут быть прерваны.
Вторая теория предполагает, что для зрительной системы мир стабилен, если не предоставлено существенных противоречивых доказательств, таких как повторное появление цели при условии отсутствия эффекта гашения (без внедрения временного пробела).
Авторы рассмотренного нами сегодня труда считают, что обе теории имеют право на существование. Даже больше, они не взаимоисключают друг друга, как показали результаты опытов и моделирования.
Опыты и моделирование также показали, что каждый из нейронных путей, затронутых в исследовании, играет свою роль в составлении визуальной картины того, что мы видим. М-путь отвечает за обнаружение смещения объекта, а Р-путь подавляет ошибочную информацию, вызванную саккадами. Таким образом, эффект SSD (саккадное подавление смещения) вступает в силу, когда сигнал Р-пути подавляет сигнал М-пути сразу после саккады. В противном случае информация содержала бы неточности (утрировано говоря, представьте себе ситуацию, когда человек двигается во время фотосъемки).
Зрительная система не только крайне важна для человека, поскольку позволяет получать большую часть информации об окружающем мире, но и крайне сложна. Наши глаза постоянно получают новую информацию. Даже если мы будем смотреть на один и тот же объект, наша сетчатка продолжает двигаться (саккады), собирая данные об объекте. И за счет того, что одни подсистемы собирают данные, а другие отфильтровывают ненужные и ложные, мы видим мир таким, каков он есть.
В дальнейшем авторы исследования намерены продолжить свою работу, поскольку предстоит еще очень во многом разобраться, прежде чем заявлять о том, что мы на 100% знаем, как работает зрительная система человека.
Благодарю за внимание, оставайтесь любопытствующими и хорошей всем рабочей недели, ребята. :)
Немного рекламы
Спасибо, что остаётесь с нами. Вам нравятся наши статьи? Хотите видеть больше интересных материалов? Поддержите нас, оформив заказ или порекомендовав знакомым, облачные VPS для разработчиков от $4.99, уникальный аналог entry-level серверов, который был придуман нами для Вас: Вся правда о VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps от $19 или как правильно делить сервер? (доступны варианты с RAID1 и RAID10, до 24 ядер и до 40GB DDR4).
Dell R730xd в 2 раза дешевле в дата-центре Equinix Tier IV в Амстердаме? Только у нас 2 х Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 ТВ от $199 в Нидерландах! Dell R420 — 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB — от $99! Читайте о том Как построить инфраструктуру корп. класса c применением серверов Dell R730xd Е5-2650 v4 стоимостью 9000 евро за копейки?


