
Несколько недель назад, после прогулки по лесам, я написал статью загадке разнообразия лесных деревьев. На своём пути я нашёл с десяток видов, имеющих общую среду обитания и, очевидно, конкурирующих за одинаковые ресурсы — в основном, за доступ к солнечному свету. Экологический принцип гласит, что один вид должен выиграть в этой конкуренции и вытеснить все остальные, но деревья, похоже, не читали учебников по экологии.
В своём эссе я также упомянул ещё три беспокоивших меня вопроса о деревьях. В статье-продолжении я хочу чуть глубже изучить эти три вопроса.

Сверху: белый дуб (Quercus alba), красный дуб (Quercus rubra), горный дуб (Quercus palustris), сахарный клён (Acer saccharum). Внизу: кария овальная (Carya ovata), берёза вишнёвая (Betula lenta). Все образцы были собраны вдоль тропы Роберта Фроста в Амхерсте, штат Массачусетс, с деревьев, находящихся на расстоянии не более ста метров друг от друга.
У ботаников есть сложный терминологический словарь для описания формы листьев: сердцевидный (как сердце на валентинке), клиновидный, мечевидный, стреловидный, копьевидный, обратноланцетный (перевёрнутый копьевидный), пальчатый (листья расходятся как пальцы), гитарообразный, почкообразный, крупнопильчатый, лопатообразный. И этот список ни в коем случае не исчерпывающий.
Стивен Вогел в своей книге The Life of a Leaf 2012 года перечисляет множество факторов и сил, которые могут иметь влияние на форму листа. Например, листья не могут быть слишком тяжёлыми, иначе они сломают черешок. С другой стороны, они не могут быть слишком хрупкими и тонкими, иначе их разорвёт ветер. Кроме того, листья не должны создавать слишком большое аэродинамическое сопротивление, иначе во время бури всё дерево может упасть.
Основная задача листа — фотосинтез: получение солнечного света, соединение молекул углекислого газа и воды, синтез углеводов. Эффективное выполнение этой задачи накладыает дополнительные ограничения на структуру листа. По мере возможности лист должен поворачивать свою поверхность к солнцу, максимизируя поток поглощаемых фотонов. Но важен и контроль температуры; биосинтезирующая система прекращает свою работу, если лист слишком горячий или холодный.
Вогел указывает, что малозаметные особенности формы листа могут иметь значимое влияние на тепловые и аэродинамические характеристики. Например, конвекционное охлаждение наиболее эффективно по краям листа; температура растёт с увеличением расстояния до ближайшего края. В средах, где существует риск перегрева, формы листьев минимизируют это расстояние; например, похоже, что лопастные формы дубовых листьев будут иметь преимущество над более простыми, округлыми формами. Но выбор между вычурными и компактными формами зависит и от других факторов. Широкие листья с выпуклыми формами получают больше всего солнечного света, но это не всегда может быть хорошо. Листья с кружевовидной структурой позволяют пятнам солнечного света проникать дальше и участвовать в фотосинтезе нескольким слоям листьев.
Естественный отбор — превосходный инструмент для подбора компромиссов между этих взаимосвязанных критериев. Если существует одна комбинация признаков, лучше всего подходящая для листьев, растущих в определённой среде обитания, то можно ожидать, что эволюция её найдёт. Но я не наблюдаю свидетельств схождения к оптимальному решению. Напротив, даже близко связанные виды имеют довольно сильно отличающиеся листья.
Взгляните на листья трёх дубов в левом верхнем углу представленной выше фотографии. Очевидно, что они являются различными вариациями одной темы. Общим для этих листьев является последовательность напоминающих полуострова выступов, ответвляющихся по очереди слева и справа от центральной линии. Вариации заключаются в количестве выступов (в этих образцах — от трёх до пяти на каждую сторону), их форме (закруглённые или острые) и глубине впадин между «полуостровами». Причинами таких вариаций могут быть генетические различия всего в нескольких локусах. Но почему листья приобрели такие различные характеристики? Из-за какой эволюционной силы для белого дуба лучше скруглённые края, а для красного и горного — острые?
Учёные многое узнали о механизмах развития, генерирующих форму листьев. С точки зрения биохимии основными действующими силами являются растительные гормоны, называемые ауксинами; их пространственное распределение и перемещение по тканям растения регулируют локальные скорости роста, а значит, и паттерн развития. (В обзорной статье 2014 года, написанной Джереми Дхаром и Ашвани Париком, подробно описываются эти аспекты формы листьев.) В математике Адам Рунионс, Милтос Тсиантис и Пшемыслав Прусинкевич вывели алгоритм, способный генерировать широкий диапазон форм листьев с поразительной похожестью на реальные образцы. (Их статья 2017 года вместе с исходным кодом и видео выложены по адресу algorithmicbotany.org/papers/leaves2017.html.) При разных значениях параметров одна и та же программа генерирует формы, которые можно распознать как дубы, клёны, платаны и другие деревья. Однако все эти работы отвечают на вопрос как, а не зачем.
Похоже, что ещё одно свойство листьев деревьев — их размер — реагирует на давление эволюции очень простым способом. Среди всех наземных растений (не только деревьев) площадь листа варьируется на порядок миллиона — от примерно 1 кв. миллиметра на лист до 1 кв. метра. В своей статье 2017 года Иэн Райт и его коллеги отмечают, что эта вариативность сильно коррелирует с климатом. В тёплых влажных регионах предпочтение отдаётся большим листьям; вспомним, например, банан. В холодных сухих средах, например, на альпийских хребтах, в основном растут крошечные растения с ещё более мелкими листьями. То есть естественный отбор в сфере листьев деревьев вполне процветает; нам просто кажется, что чётких предпочтений относительно формы листьев нет.
Или я упускаю что-то важное? Кое-где в природе мы находим вычурные вариации, которые кажутся неоправданными, если рассматривать их только с мрачной точки зрения выживания наиболее приспособленного. Подумайте, например, о красочной раскраске перьев птиц. На заднем дворе моего дома частыми гостями становятся кардиналы и голубые сойки, но я не трачу время на размышления о том, красный или синий цвет оптимальнее для выживания в этой среде обитания. Как и не ожидаю я, что эти два вида сольются в один, имеющий какой-нибудь оттенок фиолетового. Их броское оперение является не адаптацией к физической среде, а элементами системы коммуникаций; они отправляют сигналы конкурентам или потенциальным половым партнёрам. Может ли что-то подобное происходит с формой листа? Сохраняют ли разные виды дуба уникальные листья, чтобы демонстрировать себя животным, помогающим с опылением или распространением семян? Я считаю эту мысль маловероятной, но ничего лучше у меня нет.
Лесные деревья Востока США часто вырастают до 25 или 30 метров (около 100 футов). Для возведения подобной конструкции требуются огромные инвестиции материала и энергии. Для обеспечения достаточной прочности и жёсткости обхват ствола должен расти как степень от высоты, а значит, площадь его сечения растёт как куб от высоты. Из этого следует, что удвоение высоты дерева увеличивает его массу в 16 раз.
Большая высота создаёт необходимость ещё одних (постоянных) затрат — метаболических. Каждый день живое дерево должно поднимать 500 литров воды (массой 500 кг) от зоны корней на уровне земли до листьев кроны. Это подобно перетаскиванию воды, достаточной для заполнения четырёх-пяти ванн, из подвала здания на десятый этаж.
Также высота увеличивает угрозы жизни и здоровью дерева. Более высокий ствол создаёт более длинный рычаг для любой силы, которая может и свалить дерево. В дополнение к этому риску, средние скорости ветра растут с увеличением расстояния до земли.

Стоя на лесной почве, я запрокидываю голову и смотрю на лиственные кроны, находящиеся сверху огромных деревянных колонн. Я не могу не считать эти деревья на подпорках огромными пустыми тратами ресурсов. Они выглядят ещё глупее, чем иглоподобные башни апартаментов для миллиардеров, пронизывающие сегодня очертания Манхэттена. В таких зданиях все этажи имеют хоть какое-то применение. В лесу стволы деревьев оголяются от листьев, а иногда и от ветвей на 90 процентов от своей длины; здесь заняты только пентхаусы.
Если бы деревья каким-то образом могли собраться вместе и заключить сделку — установить порядок зонирования или кодекс строительства, то от этого бы выиграли они все. Возможно, они могли бы объявить максимумом высоту 10 метров. Для крон деревьев ничего бы не изменилось; это правило бы просто «урезало» нижние 20 метров ствола.
Если каждое дерево выиграло бы от такого соглашения, почему в природе эволюционно не возникают подобные «урезанные» леса? Обычно на вопрос «почему все не могут жить в согласии» отвечают, что эволюция работает не так. Общепринято, что естественный отбор является совершенно эгоистичным и индивидуалистским, даже если это вредит. Достигшее предела высоты в 10 метров дерево говорит себе: «Да, это хорошо: я получаю достаточно света без необходимости стоять на цыпочках. Но ведь может быть и ещё лучше – если я вытяну свой ствол ещё на один-два метра, то получу ещё большую долю солнечной энергии». Разумеется, другие деревья рассуждают точно так же, поэтому эта тщетная гонка вооружений продолжается. Как сказал Вогел, предвидение явно не является сильной стороной эволюции.
Я склонен согласиться с этим суровым взглядом на эволюцию, но не совсем уверен, что он на самом деле объясняет то, что мы видим в лесу. Если в подобных ситуациях у эволюции нет места совместным действиям, как получается, что все деревья прекращают рост примерно на одной высоте? В частности, если соглашение о пределе роста в 10 метров было бы нарушено неистовым жульничеством, то почему же то же самое не происходит на 30 метрах?
Можно предположить, что 30 метров являются физиологическим пределом, что деревья расли бы выше, если бы могли, но этому мешает какое-то физическое ограничение. Вероятно, они не могут поднимать воду выше. Я бы посчитал это очень многообещающей гипотезой, если бы не секвойи и калифорнийские мамонтовые деревья на Западе США. Эти деревья не слышали ни о каких подобных физических барьерах. Они спокойно вырастают до 70 или 80 метров, а некоторые образцы превышают 100 метров. Следовательно мы должны задать деревьям Западного побережья не только вопрос «Почему вы такие высокие?», но и «Почему вы не выше?»
Я могу придумать по крайней мере одну убедительную причину, по которой лесные деревья могут расти до одинаковой высоты. Если дерево ниже среднего, то оно будет страдать от нахождения в тени. Но высовывание головы и плеч из толпы тоже имеет свои недостатки: такое выделяющееся дерево будет подвергаться более сильным ветрам, большему весу льда и снега, а также, вероятно, иметь повышенную вероятность удара молнии. То есть слишком сильное отклонение в любую сторону от средней высоты может быть наказано снижением репродуктивного успеха. Однако остаётся важный вопрос: как все деревья достигают консенсуса в том, какая высота лучше?
Ещё один возможный вариант: может быть, высота лесных деревьев является не результатом гонки вооружений, а реакцией на хищников. Деревья поднимают свои листья вверх, чтобы спасти их от травоядных. Не могу сказать, что это не так, но мне кажется это маловероятным. В лесах Северной Америки не бродят жирафы (а если бы и бродили, то 10 метров было бы больше чем достаточно, чтобы защитить листья). Большинство поедающих листья деревьев животных – это членистоногие, которые или умеют летать (взрослые насекомые), или ползают по стволу (гусеницы и другие личинки). Следовательно, высота не может полностью защитить листья, и в лучшем случае способна служить лишь сдерживающим фактором. Листья деревьев – неполноценное питание; вероятно, некоторые мелкие травоядные посчитают, что за ними стоит взбираться на 10 метров, но не на 30.
На представленной ниже схеме показано два математических графа. Это набор точек (более формально называемых вершинами), соединённый отрезками прямых (рёбрами).
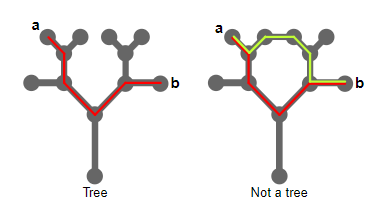
Говорят, что граф связанный, если можно перейти от любой вершины к любой другой вершине по определённой последовательности рёбер. Оба показанных выше графа являются связанными. Математические деревья являются подвидом связанных графов. Они минимально связаны: между любыми двумя вершинами есть ровно один путь.
[Тонкая техническая деталь: путь – это последовательность рёбер, в каждой любое ребро может встречаться не более одного раза. Это исключает бессмысленные возвраты туда-обратно. Последовательность вида x, y, x, z не является путём.]
Граф слева является деревом. Красной линией показан уникальный путь от a к b. Граф справа не является деревом. В нём есть два пути от a в b (красная и жёлтая линии).
Вот ещё один способ описания математического дерева: это граф, подчиняющийся антиматримониальному правилу — когда ветви расходятся в стороны, ни одна из них не соединяется снова. Биологические деревья в общем случае устроены так же: две ответвившихся от ствола ветки не вернутся к стволу и не объединятся друг с другом. Другими словами, в них нет циклов или замкнутых петель. Паттерн расходящихся ветвей, которые никогда не сойдутся вместе, очевидно заметен в повторяющейся структуре показанного ниже дерева. (Это дерево — сосна с острова Норфолк, аборигенная для южной части Тихого океана, но этот образец был сфотографирован в Сардинии.)

Деревья достигли больших успехов и без петель в ветвях. Зачем растению вообще может понадобиться, чтобы его структурные элементы росли кругами?
Я могу придумать две причины. Первая — это механическая прочность и устойчивость. Инженеры знают насколько ценны треугольники (минимально возможные замкнутые петли) в проектировании жёстких конструкций. Как ценны и арки — два вертикальных элемента, которые не могут стоять по отдельности. не опираясь друг на друга. Деревья не могут воспользоваться преимуществами подобных трюков; их ветви являются консолями, поддерживаемыми только в точке соединения со стволом или родительской веткой. Структуры с петлями позволили бы создавать различные виды распорок и креплений. Вторая причина — надёжность. Наличие множественных каналов от корней к листьям повысило бы надёжность системы циркуляции дерева. Травма рядом с основанием ветви больше бы не обрекала на смерть все структуры выше точки повреждения.
Сети с множественными путями между узлами применяются в других областях природы, и даже в других аспектах анатомии деревьев. Сетчатые каналы на показанном ниже изображении — это жилки, распределяющие жидкости и питательные вещества в листе красного дуба. Самые крупные жилы имеют древообразную структуру, однако каналы поменьше образуют вложенную иерархию петель внутри петель. (Паттерн напоминает мне карту старинного города.) Благодаря множеству избыточных путей насекомое, откусившее часть посередине этой сети, не перекроет коммуникации с остальной частью листа.

Отсутствие петель в структурах большего размера (стволах и ветвях) может быть естественным последствием программы развития, управляющей ростом дерева. Для описания такого роста венгро-голландский биолог Аристид Линдермайер изобрёл семейство формальных языков (называемых теперь L-системами). Языки — это перезаписываемые системы: мы начинаем с одного символа (аксиомы) и заменяем её на строку символов, определяемую правилами грамматики. Затем строка, полученная от этой замены, становится новыми входными данными того же процесса перезаписи, а каждый из её символов заменяется другой строкой, образованной согласно правилам грамматики. В конце символы интерпретируются как команды для построения геометрической фигуры.
Вот пример грамматики L-системы для построения мультяшных двухмерных деревьев:
Символы
Начав с аксиомы
Когда этот процесс перезаписи продолжается ещё в течение нескольких этапов, а затем преобразуется в графический вывод, то мы видим побег, вырастающий в молодое дерево, форма которого напоминает вяз.
[Я упустил некоторые детали. На каждом этапе длина шага forward (вперёд) уменьшается на коэффициент 0,6. А все повороты, как влево (left), так и вправо (right) выполняются на угол 20 градусов.]

Подобные L-системы могут создавать широкий диапазон различных ветвящихся структур. Более сложные версии такой же программы способны создавать реалистичные изображения биологических деревьев. (На веб-сайте Algorithmic Botany Университета Калгари есть множество примеров.) Чего L-системы не могут, так это создавать замкнутые петли. Для этого потребовался бы фундаментально иной тип грамматики, например, правило трансформации, получающее два символа или строки и создающее соединённый результат. (Обратите внимание, что на этапе 5 показанной выше схемы две ветви дерева кажутся наложенными друг на друга, но они не объединены. Граф не имеет вершины в точке пересечения.)
Если бы биомеханические механизмы, управляющие ростом и развитием деревьев, работали с теми же ограничениями, что и L-системы, у нас было бы удобное объяснение отсутствия петель в ветвлении биологических деревьев. Но, вероятно, это объяснение слишком уж удобное. Я говорил, что у деревьев нет петель, и в общем случае это так. Но как насчёт показанного ниже дерева — сфотографированной мной несколько лет назад лягерстремии на улицах Роли, штат Северная Каролина? (Она напоминает мне выразительную скульптуру Генри Мура.)

Это растение является деревом в ботаническом смысле, но совершенно точно не в математическом. Один ствол поднимается из земли и сразу же разделяется. На высоте моей талии у дерева четыре ветви, а чуть выше три из них воссоединяются. На высоте моей груди есть ещё одно разделение и ещё одно слияние. Дерево-бунтарь надсмехается над всеми канонами и традициями деревьев.
И лягерстремия является не единственным нарушителем правил. Горизонтальные ветки аборигенных для Индии баньянов подпёрты множеством доходящих до земли упоров. Показанный ниже баньян из Хило, Гавайи, в месте, где должен быть ствол, имеет пустую полость, окружённую десятками или сотнями поддерживающих побегов с перемычками над ними. Описанная выше L-система никогда бы не смогла создать такую сеть. Но если баньян способен на это, почему же другие деревья не пользуются тем же трюком?

В биологии вопрос «Почему x?» является укороченной версией «Каково эволюционное преимущество x?» или «Какой вклад вносит x в выживание и репродуктивный успех организма?» Для ответа на подобные вопросы часто нужен полёт воображения. Мы видим пятнистого коричневого мотылька на коре дерева и предполагаем, что его раскраска — это камуфляж, скрывающий насекомое от хищников. Мы видим яркую бабочку и приходим к выводу, что её обличье отпугивает, это предупреждение, сообщающее «Я ядовита, ты пожалеешь, если съешь меня».
Подобные объяснения имеют опасность превращения в простые сказки, наподобие истории Киплинга о том, как слон получил хобот в борьбе с крокодилом. [Французы и в самом деле называют их «почему-сказками» — les contes des pourquoi.]. Вместо дарвиновского механизма мутаций и отбора мы склонны думать с точки зрения потребностей и желаний отдельной личности. Это сложно, когда личность является животным, чья психическая жизнь (если она вообще есть) сильно далека от нашей. Боится ли мотылёк быть съеденным? Радуется ли бабочка, попав на полную полевых цветов солнечную лужайку? Мы этого не знаем.
И если уж нам сложно представить ощущения животных, то жизнь растений ещё дальше от нашего предела познаний. Жаждет ли цветок прилёта несущей пыльцу пчелы? Скорбит ли дуб, когда его жёлуди съедают белки? Что деревья думают о дятлах? Встретившись с подобными вопросами, я могу только пожать плечами. Я понятия не имею, чего хотят и боятся растения.
Другие люди заявляют, что знают гораздо больше о чувствах растительности. Немецкий лесник Петер Воллебен опубликовал книгу под названием «Тайная жизнь деревьев: что они чувствуют, как они общаются» (The Hidden Life of Trees: What They Feel, How They Communicate). Он сообщает, что деревья вскармливает своё потомство, поддерживают дружеские отношения с соседями и защищают болеющих и раненных членов своего сообщества. В определённой мере эти идеи имеют научное обоснование, они позаимствованы из работы, проделанной в лаборатории Сюзанн Симард в Университете Британской Колумбии. Симард, являющаяся руководителем проекта Mother Tree, изучает коммуникационные сети, образованные корнями деревьев и связанными с ними почвенными грибами.
Работа Симард кажется мне интересной. Но я считаю антропоморфическую реторику бесполезной и агрессивной. Её цель, насколько я понимаю, заключается в том, чтобы мы больше заботились о деревьях и лесах, считая, что они во многом похожи на нас; у них есть семьи и сообщества, дружба, альянсы. На мой взгляд, это совершенно неправильно. Сильнее всего деревья интригуют тем, что они являются чужаками среди нас, живыми существами, чья долгая, неподвижная и тихая жизнь никак не напоминает нашу собственную неистовую суматоху. Деревья представляют огромную загадку сами по себе, без всяких попыток их очеловечивания.
Брайан Хейес — мой любимый автор научно-популярных статей, поэтому за годы на Хабре накопилась целая серия его переводов:
Dkhar, Jeremy, and Ashwani Pareek. 2014. What determines a leaf’s shape? EvoDevo 5:47.
McMahon, Thomas A. 1975. The mechanical design of trees. Scientific American 233(1):93–102.
Osnas, Jeanne L. D., Jeremy W. Lichstein, Peter B. Reich, and Stephen W. Pacala. 2013. Global leaf trait relationships: mass, area, and the leaf economics spectrum. Science 340:741–744.
Prusinkiewicz, Przemyslaw, and Aristid Lindenmayer, with James S. Hanan, F. David Fracchia, Deborah Fowler, Martin J. M. de Boer, and Lynn Mercer. 1990. The Algorithmic Beauty of Plants. New York: Springer-Verlag. PDF edition available at http://algorithmicbotany.org/papers/.
Runions, Adam, Martin Fuhrer, Brendan Lane, Pavol Federl, Anne-Gaëlle Rolland-Lagan, and Przemyslaw Prusinkiewicz. 2005. Modeling and visualization of leaf venation patterns. ACM Transactions on Graphics 24(3):702-711.
Runions, Adam, Miltos Tsiantis, and Przemyslaw Prusinkiewicz. 2017. A common developmental program can produce diverse leaf shapes. New Phytologist 216:401–418. Preprint and source code.
Tadrist, Loïc, and Baptiste Darbois Texier. 2016. Are leaves optimally designed for self-support? An investigation on giant monocots. arXiv:1602.03353.
Vogel, Steven. 2012. The Life of a Leaf. University of Chicago Press.
Wright, Ian J., Ning Dong, Vincent Maire, I. Colin Prentice, Mark Westoby, Sandra Díaz, Rachael V. Gallagher, Bonnie F. Jacobs, Robert Kooyman, Elizabeth A. Law, Michelle R. Leishman, Ülo Niinemets, Peter B. Reich, Lawren Sack, Rafael Villar, Han Wang, and Peter Wilf. 2017. Global climatic drivers of leaf size. Science 357:917–921.
Yamazaki, Kazuo. 2011. Gone with the wind: trembling leaves may deter herbivory. Biological Journal of the Linnean Society 104:738–747.
Young, David A. 2010 preprint. Growth-algorithm model of leaf shape. arXiv:1004.4388.
В своём эссе я также упомянул ещё три беспокоивших меня вопроса о деревьях. В статье-продолжении я хочу чуть глубже изучить эти три вопроса.
Вопрос 1: Почему листья деревьев имеют такую разнообразную форму?
Сверху: белый дуб (Quercus alba), красный дуб (Quercus rubra), горный дуб (Quercus palustris), сахарный клён (Acer saccharum). Внизу: кария овальная (Carya ovata), берёза вишнёвая (Betula lenta). Все образцы были собраны вдоль тропы Роберта Фроста в Амхерсте, штат Массачусетс, с деревьев, находящихся на расстоянии не более ста метров друг от друга.
У ботаников есть сложный терминологический словарь для описания формы листьев: сердцевидный (как сердце на валентинке), клиновидный, мечевидный, стреловидный, копьевидный, обратноланцетный (перевёрнутый копьевидный), пальчатый (листья расходятся как пальцы), гитарообразный, почкообразный, крупнопильчатый, лопатообразный. И этот список ни в коем случае не исчерпывающий.
Стивен Вогел в своей книге The Life of a Leaf 2012 года перечисляет множество факторов и сил, которые могут иметь влияние на форму листа. Например, листья не могут быть слишком тяжёлыми, иначе они сломают черешок. С другой стороны, они не могут быть слишком хрупкими и тонкими, иначе их разорвёт ветер. Кроме того, листья не должны создавать слишком большое аэродинамическое сопротивление, иначе во время бури всё дерево может упасть.
Основная задача листа — фотосинтез: получение солнечного света, соединение молекул углекислого газа и воды, синтез углеводов. Эффективное выполнение этой задачи накладыает дополнительные ограничения на структуру листа. По мере возможности лист должен поворачивать свою поверхность к солнцу, максимизируя поток поглощаемых фотонов. Но важен и контроль температуры; биосинтезирующая система прекращает свою работу, если лист слишком горячий или холодный.
Вогел указывает, что малозаметные особенности формы листа могут иметь значимое влияние на тепловые и аэродинамические характеристики. Например, конвекционное охлаждение наиболее эффективно по краям листа; температура растёт с увеличением расстояния до ближайшего края. В средах, где существует риск перегрева, формы листьев минимизируют это расстояние; например, похоже, что лопастные формы дубовых листьев будут иметь преимущество над более простыми, округлыми формами. Но выбор между вычурными и компактными формами зависит и от других факторов. Широкие листья с выпуклыми формами получают больше всего солнечного света, но это не всегда может быть хорошо. Листья с кружевовидной структурой позволяют пятнам солнечного света проникать дальше и участвовать в фотосинтезе нескольким слоям листьев.
Естественный отбор — превосходный инструмент для подбора компромиссов между этих взаимосвязанных критериев. Если существует одна комбинация признаков, лучше всего подходящая для листьев, растущих в определённой среде обитания, то можно ожидать, что эволюция её найдёт. Но я не наблюдаю свидетельств схождения к оптимальному решению. Напротив, даже близко связанные виды имеют довольно сильно отличающиеся листья.
Взгляните на листья трёх дубов в левом верхнем углу представленной выше фотографии. Очевидно, что они являются различными вариациями одной темы. Общим для этих листьев является последовательность напоминающих полуострова выступов, ответвляющихся по очереди слева и справа от центральной линии. Вариации заключаются в количестве выступов (в этих образцах — от трёх до пяти на каждую сторону), их форме (закруглённые или острые) и глубине впадин между «полуостровами». Причинами таких вариаций могут быть генетические различия всего в нескольких локусах. Но почему листья приобрели такие различные характеристики? Из-за какой эволюционной силы для белого дуба лучше скруглённые края, а для красного и горного — острые?
Учёные многое узнали о механизмах развития, генерирующих форму листьев. С точки зрения биохимии основными действующими силами являются растительные гормоны, называемые ауксинами; их пространственное распределение и перемещение по тканям растения регулируют локальные скорости роста, а значит, и паттерн развития. (В обзорной статье 2014 года, написанной Джереми Дхаром и Ашвани Париком, подробно описываются эти аспекты формы листьев.) В математике Адам Рунионс, Милтос Тсиантис и Пшемыслав Прусинкевич вывели алгоритм, способный генерировать широкий диапазон форм листьев с поразительной похожестью на реальные образцы. (Их статья 2017 года вместе с исходным кодом и видео выложены по адресу algorithmicbotany.org/papers/leaves2017.html.) При разных значениях параметров одна и та же программа генерирует формы, которые можно распознать как дубы, клёны, платаны и другие деревья. Однако все эти работы отвечают на вопрос как, а не зачем.
Похоже, что ещё одно свойство листьев деревьев — их размер — реагирует на давление эволюции очень простым способом. Среди всех наземных растений (не только деревьев) площадь листа варьируется на порядок миллиона — от примерно 1 кв. миллиметра на лист до 1 кв. метра. В своей статье 2017 года Иэн Райт и его коллеги отмечают, что эта вариативность сильно коррелирует с климатом. В тёплых влажных регионах предпочтение отдаётся большим листьям; вспомним, например, банан. В холодных сухих средах, например, на альпийских хребтах, в основном растут крошечные растения с ещё более мелкими листьями. То есть естественный отбор в сфере листьев деревьев вполне процветает; нам просто кажется, что чётких предпочтений относительно формы листьев нет.
Или я упускаю что-то важное? Кое-где в природе мы находим вычурные вариации, которые кажутся неоправданными, если рассматривать их только с мрачной точки зрения выживания наиболее приспособленного. Подумайте, например, о красочной раскраске перьев птиц. На заднем дворе моего дома частыми гостями становятся кардиналы и голубые сойки, но я не трачу время на размышления о том, красный или синий цвет оптимальнее для выживания в этой среде обитания. Как и не ожидаю я, что эти два вида сольются в один, имеющий какой-нибудь оттенок фиолетового. Их броское оперение является не адаптацией к физической среде, а элементами системы коммуникаций; они отправляют сигналы конкурентам или потенциальным половым партнёрам. Может ли что-то подобное происходит с формой листа? Сохраняют ли разные виды дуба уникальные листья, чтобы демонстрировать себя животным, помогающим с опылением или распространением семян? Я считаю эту мысль маловероятной, но ничего лучше у меня нет.
Вопрос 2: Почему деревья такие высокие?Да, этот вопрос слишком прост! Мы знаем, почему деревья растут ввысь — они стремятся к небу. Это их единственная надежда сбежать из сумрачных глубин нижних ярусов леса и получить свою долю солнечных лучей. Другими словами, если ты лесное дерево, тебе нужно расти ввысь, потому что высоки твои соседи: они закрывают тебя тенью. А соседи растут ввысь, потому что высок ты. Это классическая гонка вооружений. У Вогела есть проницательный комментарий на эту тему:
В любом ряду поколений, независимо пришедших к форме древоподобных растений, множество видов достигает больших высот. Я воспринимаю это как высоту глупости… Мы с большой вероятностью видим живой пример ограничений эволюционного планирования.Разумеется, порицание Вогелом дарвиновкой эволюции — всего лишь шутка. Однако я считаю, что вопрос договоров об ограничении высоты заслуживает более серьёзного внимания.
Договорённость об ограничении размеров ствола позволила бы всем особям приносить больше семян и начинать выращивать семена в более раннем возрасте. Однако глупый процесс эволюции не смог этого понять — предвидение явно не является его сильной стороной.
Лесные деревья Востока США часто вырастают до 25 или 30 метров (около 100 футов). Для возведения подобной конструкции требуются огромные инвестиции материала и энергии. Для обеспечения достаточной прочности и жёсткости обхват ствола должен расти как степень от высоты, а значит, площадь его сечения растёт как куб от высоты. Из этого следует, что удвоение высоты дерева увеличивает его массу в 16 раз.
Большая высота создаёт необходимость ещё одних (постоянных) затрат — метаболических. Каждый день живое дерево должно поднимать 500 литров воды (массой 500 кг) от зоны корней на уровне земли до листьев кроны. Это подобно перетаскиванию воды, достаточной для заполнения четырёх-пяти ванн, из подвала здания на десятый этаж.
Также высота увеличивает угрозы жизни и здоровью дерева. Более высокий ствол создаёт более длинный рычаг для любой силы, которая может и свалить дерево. В дополнение к этому риску, средние скорости ветра растут с увеличением расстояния до земли.
Стоя на лесной почве, я запрокидываю голову и смотрю на лиственные кроны, находящиеся сверху огромных деревянных колонн. Я не могу не считать эти деревья на подпорках огромными пустыми тратами ресурсов. Они выглядят ещё глупее, чем иглоподобные башни апартаментов для миллиардеров, пронизывающие сегодня очертания Манхэттена. В таких зданиях все этажи имеют хоть какое-то применение. В лесу стволы деревьев оголяются от листьев, а иногда и от ветвей на 90 процентов от своей длины; здесь заняты только пентхаусы.
Если бы деревья каким-то образом могли собраться вместе и заключить сделку — установить порядок зонирования или кодекс строительства, то от этого бы выиграли они все. Возможно, они могли бы объявить максимумом высоту 10 метров. Для крон деревьев ничего бы не изменилось; это правило бы просто «урезало» нижние 20 метров ствола.
Если каждое дерево выиграло бы от такого соглашения, почему в природе эволюционно не возникают подобные «урезанные» леса? Обычно на вопрос «почему все не могут жить в согласии» отвечают, что эволюция работает не так. Общепринято, что естественный отбор является совершенно эгоистичным и индивидуалистским, даже если это вредит. Достигшее предела высоты в 10 метров дерево говорит себе: «Да, это хорошо: я получаю достаточно света без необходимости стоять на цыпочках. Но ведь может быть и ещё лучше – если я вытяну свой ствол ещё на один-два метра, то получу ещё большую долю солнечной энергии». Разумеется, другие деревья рассуждают точно так же, поэтому эта тщетная гонка вооружений продолжается. Как сказал Вогел, предвидение явно не является сильной стороной эволюции.
Я склонен согласиться с этим суровым взглядом на эволюцию, но не совсем уверен, что он на самом деле объясняет то, что мы видим в лесу. Если в подобных ситуациях у эволюции нет места совместным действиям, как получается, что все деревья прекращают рост примерно на одной высоте? В частности, если соглашение о пределе роста в 10 метров было бы нарушено неистовым жульничеством, то почему же то же самое не происходит на 30 метрах?
Можно предположить, что 30 метров являются физиологическим пределом, что деревья расли бы выше, если бы могли, но этому мешает какое-то физическое ограничение. Вероятно, они не могут поднимать воду выше. Я бы посчитал это очень многообещающей гипотезой, если бы не секвойи и калифорнийские мамонтовые деревья на Западе США. Эти деревья не слышали ни о каких подобных физических барьерах. Они спокойно вырастают до 70 или 80 метров, а некоторые образцы превышают 100 метров. Следовательно мы должны задать деревьям Западного побережья не только вопрос «Почему вы такие высокие?», но и «Почему вы не выше?»
Я могу придумать по крайней мере одну убедительную причину, по которой лесные деревья могут расти до одинаковой высоты. Если дерево ниже среднего, то оно будет страдать от нахождения в тени. Но высовывание головы и плеч из толпы тоже имеет свои недостатки: такое выделяющееся дерево будет подвергаться более сильным ветрам, большему весу льда и снега, а также, вероятно, иметь повышенную вероятность удара молнии. То есть слишком сильное отклонение в любую сторону от средней высоты может быть наказано снижением репродуктивного успеха. Однако остаётся важный вопрос: как все деревья достигают консенсуса в том, какая высота лучше?
Ещё один возможный вариант: может быть, высота лесных деревьев является не результатом гонки вооружений, а реакцией на хищников. Деревья поднимают свои листья вверх, чтобы спасти их от травоядных. Не могу сказать, что это не так, но мне кажется это маловероятным. В лесах Северной Америки не бродят жирафы (а если бы и бродили, то 10 метров было бы больше чем достаточно, чтобы защитить листья). Большинство поедающих листья деревьев животных – это членистоногие, которые или умеют летать (взрослые насекомые), или ползают по стволу (гусеницы и другие личинки). Следовательно, высота не может полностью защитить листья, и в лучшем случае способна служить лишь сдерживающим фактором. Листья деревьев – неполноценное питание; вероятно, некоторые мелкие травоядные посчитают, что за ними стоит взбираться на 10 метров, но не на 30.
Вопрос 3: Почему деревья представляют собой деревья (а не другие виды графов)?Для биолога дерево – это древесное растение достаточной высоты. Для математика дерево – это граф без петель. Оказывается, математические и биологические деревья имеют общие важные свойства.
На представленной ниже схеме показано два математических графа. Это набор точек (более формально называемых вершинами), соединённый отрезками прямых (рёбрами).
Говорят, что граф связанный, если можно перейти от любой вершины к любой другой вершине по определённой последовательности рёбер. Оба показанных выше графа являются связанными. Математические деревья являются подвидом связанных графов. Они минимально связаны: между любыми двумя вершинами есть ровно один путь.
[Тонкая техническая деталь: путь – это последовательность рёбер, в каждой любое ребро может встречаться не более одного раза. Это исключает бессмысленные возвраты туда-обратно. Последовательность вида x, y, x, z не является путём.]
Граф слева является деревом. Красной линией показан уникальный путь от a к b. Граф справа не является деревом. В нём есть два пути от a в b (красная и жёлтая линии).
Вот ещё один способ описания математического дерева: это граф, подчиняющийся антиматримониальному правилу — когда ветви расходятся в стороны, ни одна из них не соединяется снова. Биологические деревья в общем случае устроены так же: две ответвившихся от ствола ветки не вернутся к стволу и не объединятся друг с другом. Другими словами, в них нет циклов или замкнутых петель. Паттерн расходящихся ветвей, которые никогда не сойдутся вместе, очевидно заметен в повторяющейся структуре показанного ниже дерева. (Это дерево — сосна с острова Норфолк, аборигенная для южной части Тихого океана, но этот образец был сфотографирован в Сардинии.)
Деревья достигли больших успехов и без петель в ветвях. Зачем растению вообще может понадобиться, чтобы его структурные элементы росли кругами?
Я могу придумать две причины. Первая — это механическая прочность и устойчивость. Инженеры знают насколько ценны треугольники (минимально возможные замкнутые петли) в проектировании жёстких конструкций. Как ценны и арки — два вертикальных элемента, которые не могут стоять по отдельности. не опираясь друг на друга. Деревья не могут воспользоваться преимуществами подобных трюков; их ветви являются консолями, поддерживаемыми только в точке соединения со стволом или родительской веткой. Структуры с петлями позволили бы создавать различные виды распорок и креплений. Вторая причина — надёжность. Наличие множественных каналов от корней к листьям повысило бы надёжность системы циркуляции дерева. Травма рядом с основанием ветви больше бы не обрекала на смерть все структуры выше точки повреждения.
Сети с множественными путями между узлами применяются в других областях природы, и даже в других аспектах анатомии деревьев. Сетчатые каналы на показанном ниже изображении — это жилки, распределяющие жидкости и питательные вещества в листе красного дуба. Самые крупные жилы имеют древообразную структуру, однако каналы поменьше образуют вложенную иерархию петель внутри петель. (Паттерн напоминает мне карту старинного города.) Благодаря множеству избыточных путей насекомое, откусившее часть посередине этой сети, не перекроет коммуникации с остальной частью листа.
Отсутствие петель в структурах большего размера (стволах и ветвях) может быть естественным последствием программы развития, управляющей ростом дерева. Для описания такого роста венгро-голландский биолог Аристид Линдермайер изобрёл семейство формальных языков (называемых теперь L-системами). Языки — это перезаписываемые системы: мы начинаем с одного символа (аксиомы) и заменяем её на строку символов, определяемую правилами грамматики. Затем строка, полученная от этой замены, становится новыми входными данными того же процесса перезаписи, а каждый из её символов заменяется другой строкой, образованной согласно правилам грамматики. В конце символы интерпретируются как команды для построения геометрической фигуры.
Вот пример грамматики L-системы для построения мультяшных двухмерных деревьев:
f ⟶ f [r f] [l f]
l ⟶ l
r ⟶ rСимволы
f, l и r являются базовыми элементами языка; в процессе интерпретации в качестве команд для рисования, они обозначают forward («вперёд»), left («влево») и right («вправо»). Первое правило грамматики заменяет любое вхождение f строкой f [l f] [r f]; второе и третье правила ничего не меняют, заменяя l и r на них же. Квадратные скобки являются границами подпрограммы. Дойдя до левой скобки, система запоминает свою текущую позицию и ориентацию рисунка. Затем она исполняет инструкции внутри скобок, а дойдя до правой скобки, возвращается назад к сохранённой позиции и ориентации.Начав с аксиомы
f, грамматика создаёт последовательность ещё более сложных последовательностей команд:Этап 0: f Этап 1: f [r f] [l f] Этап 2: f [r f] [l f] [r f [r f] [l f]] [l f [r f] [l f]]]
Когда этот процесс перезаписи продолжается ещё в течение нескольких этапов, а затем преобразуется в графический вывод, то мы видим побег, вырастающий в молодое дерево, форма которого напоминает вяз.
[Я упустил некоторые детали. На каждом этапе длина шага forward (вперёд) уменьшается на коэффициент 0,6. А все повороты, как влево (left), так и вправо (right) выполняются на угол 20 градусов.]
Подобные L-системы могут создавать широкий диапазон различных ветвящихся структур. Более сложные версии такой же программы способны создавать реалистичные изображения биологических деревьев. (На веб-сайте Algorithmic Botany Университета Калгари есть множество примеров.) Чего L-системы не могут, так это создавать замкнутые петли. Для этого потребовался бы фундаментально иной тип грамматики, например, правило трансформации, получающее два символа или строки и создающее соединённый результат. (Обратите внимание, что на этапе 5 показанной выше схемы две ветви дерева кажутся наложенными друг на друга, но они не объединены. Граф не имеет вершины в точке пересечения.)
Если бы биомеханические механизмы, управляющие ростом и развитием деревьев, работали с теми же ограничениями, что и L-системы, у нас было бы удобное объяснение отсутствия петель в ветвлении биологических деревьев. Но, вероятно, это объяснение слишком уж удобное. Я говорил, что у деревьев нет петель, и в общем случае это так. Но как насчёт показанного ниже дерева — сфотографированной мной несколько лет назад лягерстремии на улицах Роли, штат Северная Каролина? (Она напоминает мне выразительную скульптуру Генри Мура.)
Это растение является деревом в ботаническом смысле, но совершенно точно не в математическом. Один ствол поднимается из земли и сразу же разделяется. На высоте моей талии у дерева четыре ветви, а чуть выше три из них воссоединяются. На высоте моей груди есть ещё одно разделение и ещё одно слияние. Дерево-бунтарь надсмехается над всеми канонами и традициями деревьев.
И лягерстремия является не единственным нарушителем правил. Горизонтальные ветки аборигенных для Индии баньянов подпёрты множеством доходящих до земли упоров. Показанный ниже баньян из Хило, Гавайи, в месте, где должен быть ствол, имеет пустую полость, окружённую десятками или сотнями поддерживающих побегов с перемычками над ними. Описанная выше L-система никогда бы не смогла создать такую сеть. Но если баньян способен на это, почему же другие деревья не пользуются тем же трюком?
В биологии вопрос «Почему x?» является укороченной версией «Каково эволюционное преимущество x?» или «Какой вклад вносит x в выживание и репродуктивный успех организма?» Для ответа на подобные вопросы часто нужен полёт воображения. Мы видим пятнистого коричневого мотылька на коре дерева и предполагаем, что его раскраска — это камуфляж, скрывающий насекомое от хищников. Мы видим яркую бабочку и приходим к выводу, что её обличье отпугивает, это предупреждение, сообщающее «Я ядовита, ты пожалеешь, если съешь меня».
Подобные объяснения имеют опасность превращения в простые сказки, наподобие истории Киплинга о том, как слон получил хобот в борьбе с крокодилом. [Французы и в самом деле называют их «почему-сказками» — les contes des pourquoi.]. Вместо дарвиновского механизма мутаций и отбора мы склонны думать с точки зрения потребностей и желаний отдельной личности. Это сложно, когда личность является животным, чья психическая жизнь (если она вообще есть) сильно далека от нашей. Боится ли мотылёк быть съеденным? Радуется ли бабочка, попав на полную полевых цветов солнечную лужайку? Мы этого не знаем.
И если уж нам сложно представить ощущения животных, то жизнь растений ещё дальше от нашего предела познаний. Жаждет ли цветок прилёта несущей пыльцу пчелы? Скорбит ли дуб, когда его жёлуди съедают белки? Что деревья думают о дятлах? Встретившись с подобными вопросами, я могу только пожать плечами. Я понятия не имею, чего хотят и боятся растения.
Другие люди заявляют, что знают гораздо больше о чувствах растительности. Немецкий лесник Петер Воллебен опубликовал книгу под названием «Тайная жизнь деревьев: что они чувствуют, как они общаются» (The Hidden Life of Trees: What They Feel, How They Communicate). Он сообщает, что деревья вскармливает своё потомство, поддерживают дружеские отношения с соседями и защищают болеющих и раненных членов своего сообщества. В определённой мере эти идеи имеют научное обоснование, они позаимствованы из работы, проделанной в лаборатории Сюзанн Симард в Университете Британской Колумбии. Симард, являющаяся руководителем проекта Mother Tree, изучает коммуникационные сети, образованные корнями деревьев и связанными с ними почвенными грибами.
Работа Симард кажется мне интересной. Но я считаю антропоморфическую реторику бесполезной и агрессивной. Её цель, насколько я понимаю, заключается в том, чтобы мы больше заботились о деревьях и лесах, считая, что они во многом похожи на нас; у них есть семьи и сообщества, дружба, альянсы. На мой взгляд, это совершенно неправильно. Сильнее всего деревья интригуют тем, что они являются чужаками среди нас, живыми существами, чья долгая, неподвижная и тихая жизнь никак не напоминает нашу собственную неистовую суматоху. Деревья представляют огромную загадку сами по себе, без всяких попыток их очеловечивания.
Переводы других статей Брайана Хейеса на Хабре
Брайан Хейес — мой любимый автор научно-популярных статей, поэтому за годы на Хабре накопилась целая серия его переводов:
- Почему единственный вид деревьев не победил все остальные?
- Неразбериха с Boeing 737 MAX: анализ возможных причин аварий
- 1000-мерный куб: можно ли сегодня создать вычислительную модель человеческой памяти?
- Считаем кур, пока их не заклевали
- Короткое плечо совпадения
- Структура и случайность простых чисел
- Проблемы эгоистов: дорожные пробки и парадокс Браеса
- Небольшое расхождение
Дополнительное чтение
Dkhar, Jeremy, and Ashwani Pareek. 2014. What determines a leaf’s shape? EvoDevo 5:47.
McMahon, Thomas A. 1975. The mechanical design of trees. Scientific American 233(1):93–102.
Osnas, Jeanne L. D., Jeremy W. Lichstein, Peter B. Reich, and Stephen W. Pacala. 2013. Global leaf trait relationships: mass, area, and the leaf economics spectrum. Science 340:741–744.
Prusinkiewicz, Przemyslaw, and Aristid Lindenmayer, with James S. Hanan, F. David Fracchia, Deborah Fowler, Martin J. M. de Boer, and Lynn Mercer. 1990. The Algorithmic Beauty of Plants. New York: Springer-Verlag. PDF edition available at http://algorithmicbotany.org/papers/.
Runions, Adam, Martin Fuhrer, Brendan Lane, Pavol Federl, Anne-Gaëlle Rolland-Lagan, and Przemyslaw Prusinkiewicz. 2005. Modeling and visualization of leaf venation patterns. ACM Transactions on Graphics 24(3):702-711.
Runions, Adam, Miltos Tsiantis, and Przemyslaw Prusinkiewicz. 2017. A common developmental program can produce diverse leaf shapes. New Phytologist 216:401–418. Preprint and source code.
Tadrist, Loïc, and Baptiste Darbois Texier. 2016. Are leaves optimally designed for self-support? An investigation on giant monocots. arXiv:1602.03353.
Vogel, Steven. 2012. The Life of a Leaf. University of Chicago Press.
Wright, Ian J., Ning Dong, Vincent Maire, I. Colin Prentice, Mark Westoby, Sandra Díaz, Rachael V. Gallagher, Bonnie F. Jacobs, Robert Kooyman, Elizabeth A. Law, Michelle R. Leishman, Ülo Niinemets, Peter B. Reich, Lawren Sack, Rafael Villar, Han Wang, and Peter Wilf. 2017. Global climatic drivers of leaf size. Science 357:917–921.
Yamazaki, Kazuo. 2011. Gone with the wind: trembling leaves may deter herbivory. Biological Journal of the Linnean Society 104:738–747.
Young, David A. 2010 preprint. Growth-algorithm model of leaf shape. arXiv:1004.4388.



")
